Establishing and maintaining proper cell polarity are key aspects of life. One of the major goals of cell biology is to understand the molecular mechanisms mediating cell polarization. Generally, cell polarity means cellular asymmetry.
Polarization and asymmetrical growth in eukaryotic cells require elaborate regulation of a variety of cellular processes, including spatially restricted signaling, reorganization of the cytoskeleton, and polarized membrane trafficking.
Polarized cells divide asymmetrically, generating daughter cells that are different in cellular contents, cell size, or developmental potential. The asymmetric distribution of cell fate determinants (e.g. proteins and RNAs) and signaling platforms, but also asymmetric translational repression and asymmetric protein degradation control cell fate determination, cell polarity and cell differentiation, which are essential for the development of a multicellular organism or tissue containing cells with different molecular fates.
To study the establishment and maintenance of cell polarity in flowering plants we use the following Arabidopsis model cells: pollen tubes, root hairs, trichomes, and zygotes.
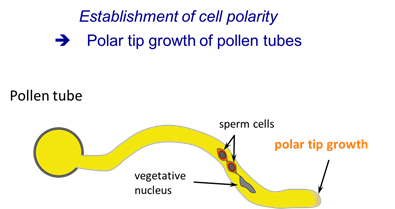
Polar tip growth of pollen tubes
The pollen tube is among the fastest growing plant cells. It is widely used as a model cell to study tip growth.
The pollen grain germinates on the surface of the stigma and forms an elongating tube that navigates through the female tissues toward the ovule to deliver its two sperm cells for double fertilization. Like growth cones of axons, pollen tubes require a signaling network to recognize and interpret extracellular cues that determine the site and the direction of tip growth. Within the past years, crucial molecular players of the pollen tube tip growth machinery have been identified, including a tip-focused calcium gradient, elaborate F-actin dynamics, tip-directed vesicle trafficking, and localized exocytosis.
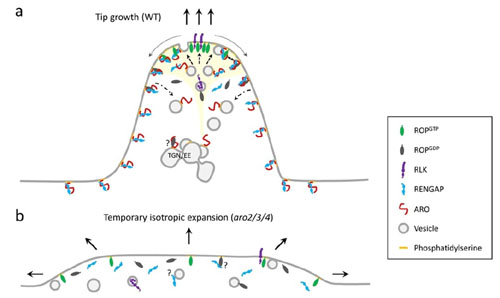
Important regulators of eukaryotic cell polarity are small GTPases of the Rho family. Rho GTPases are conserved molecular switches that cycle between a GDP-bound inactive state and a GTP-bound active state. Conformation-dependent transient interactions of Rho GTPases with regulatory and effector proteins initiate downstream responses, including changes in cytoskeletal dynamics and directional vesicle trafficking.
In our lab, we work on a small family of Armadillo repeat-containing proteins termed ARMADILLO REPEAT ONLY (ARO) proteins. The founding member, ARO1, was identified as important player for pollen tube tip growth (Gebert et al., 2008). The closely related family members ARO2, ARO3, and ARO4 are sporophyte-expressed and redundantly active in polar growing root hairs and trichomes (Kulich et al., 2020).
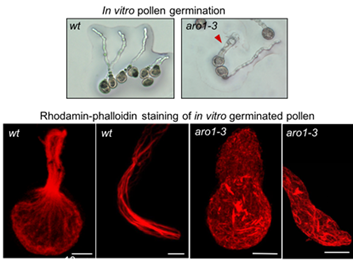
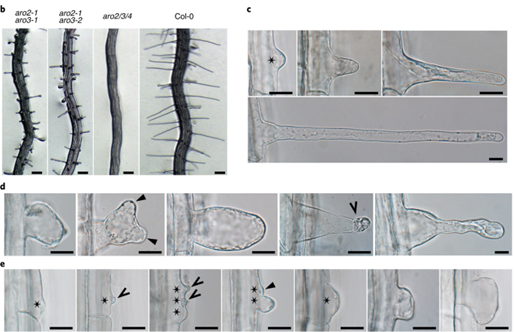
We discovered that ARO proteins contribute to the spatiotemporal regulation of ROP activity at highly polarized growth sites by recruiting GTPase-activating (GAP) domain-containing RENGAPs to these plasma membrane regions (Kulich et al., 2020). Thus, ARO proteins exhibit properties of signaling scaffolds. We are now characterizing the composition of ARO scaffolding complexes at polar growth sites and how the ARO scaffolding activity is regulated.